Carica papaya
Papaya, pawpaw



Origin
Papaya is native to Central American lowlands. It is nowadays grown in most countries within 25 degrees of the equator, with annual world production exceeding 7MT.
Climate
It is a tropical plant that nevertheless can do very well in sub-tropical climates. Ideally, temperatures should be 21-33°C and humidity above 65%. If temperature is below 12-14°C for several hours overnight, growth and productivity is compromised. Dioecious cvs are more reliable and better performers in sub-tropical climates than the hermaphrodite types. Yield is best with a minimum of 100mm per month rainfall or irrigation.
Plant Description
It is a large, rapidly-growing, tropical, herbaceous, hollow-stemmed tree, usually with a single trunk unless damaged. The large leaves are palmately lobed on long petioles. Older leaves senesce continually with most of the canopy at the apex.
Relatives
Papaya is the most important edible species in the Caricaceae family. Babaco is a small fruit-producing tree that’s a hybrid of two other species in the family.
Soils
A variety of soil types can be used provided they are well-drained. Substantial amounts of organic matter should be added to sandy soils. Preferred soil pH is 5.5-6.5.
Propagation
Almost universally by seed, as they germinate readily, and if properly dried, can be stored for lengthy periods. Seedlings may be ready for in-ground planting within several weeks of germination. Cuttings and grafting can be used but are often not worth the extra effort. Rigorous sanitation and bottom heat are advised with cuttings. Grafting can be used to transform surplus male plants into known females.
Cultivars
The best strategy for home growers is to use seed from favourable fruits produced locally, as it is then known that they perform under prevailing conditions. If the tree form can be seen in fruiting, select those where the fruit has a longish peduncle allowing it to form properly, instead of those where numerous fruit are all tightly bunched together, resulting in deformations and difficulty in picking. Retail-sourced seed could have been from fruit trees that favoured other climates.
Tropical countries generally prefer to grow hermaphrodites, and Solo is one of the better-known ones. Hawaii has authorised sale of transgenic plants to combat papaya ring spot virus which threatened the viability of the whole industry there. The genome has virtually been established.
Flowering and Pollination
Sexual behaviour in papaya is complex. In essence, there are three main types of flowers: staminate, pistillate and hermaphrodite. But in reality, there are many more variants than this, particularly amongst the hermaphrodites. Plants can be polygamo–monoecious or polygamo-dioecious. Flowers are borne on cymose inflorescences that arise from leaf axils. Pistillate flowers have an ovoid ovary with short peduncles while staminate are tubular with 10 stamens and clustered on long racemes; the hermaphroditic form is between the two.
Environmental factors (eg low temperatures, excessive N or moisture, plant damage) can cause undesirable changes in behaviour of staminate and hermaphrodite plants, whereas pistillate trees are much more stable.
Pollination is thought to be by moths; bees may be seen working staminate flowers but pistillate flowers are not attractive to them. Hermaphrodites can be pollinated before flowers open and are just as variable as open-pollinated dioecious lines, seemingly from rare but still finite cross-pollination.
Cultivation
The sex of seedlings is not known till flowering, so usually 2-3 are planted in a single hole, and once identified, the surplus can be removed by cutting them off at ground level. In reasonable proximity, one male is sufficient to pollinate several females. Space them at least 2m apart. Endeavour to keep them actively growing through all the warmer months of the year. They need a constant supply of nutrients to maintain growth and production, particularly N and K, but also micronutrients. When fruiting, they prefer high K and low P fertilization; yield starts to decrease after 3-4 years and commercial plants are usually then replaced with new stock.
Wind Tolerance
With strong winds the main damage is extensive shredding of the large leaves. However more violent storms or cyclones can break branches or even uproot the whole plant.
Pruning
They are self-pruning, with older leaves continually abscising. After a few years the plant may become too tall to access fruit easily; at this stage it can be headed back, wherafter it will regrow with several lateral branches. Although the number of fruit may then be increased, fruit size is generally smaller.
The Fruit
Papaya is a fleshy berry from 200g-5kg. Shape is sex-linked, with female flowers producing spherical-ovoid fruit and hermaphrodite more cylindrical or pear-shaped fruit. There is a central ovoid cavity normally with hundreds of black seeds. Some fruits set parthenocarpically. Male plants occasionally bear small fruits of poor quality.
The thin fruit skin changes from green to yellow-orange when mature. Flesh colour can be yellow-orange to reddish, and unless true-breeding lines are used, the red flesh types are not stable. Cut or damaged immature fruit exude a milky latex which decreases as they ripen. Fruit flavour can vary between different plants. The flesh contains 8-16% sugars with many aromatics contributing to flavour, and it also has good levels of vitamin A and C.
Fruit Production and Harvesting
Fruit can be available for much of the year in warm climates and should be harvested when about a third has changed colour. They will then ripen at room temperature over several days. As a tropical fruit they will suffer chilling injury if stored below 10°C.
Fruit Uses
Normally eaten fresh, but is also eaten green as a vegetable.
Pests and Diseases
Fruit flies. Papaya ring spot virus can destroy whole plantations, as can a phytoplasma-induced dieback when trees are under stress. Post-harvest, anthracnose can rapidly destroy whole fruit.
Comments
Along with bananas, papaya is one of the most productive fruit trees in terms of required footprint. Sourcing plants is simple and cheap via self-germinated seedlings, and with our dry WA climate, fungal infections are much reduced compared to those in the more humid tropics. Take care that soil is well-drained and maintain growth through watering and fertilizing.
More Information
Opening the Door on the Mysterious Sexual Antics of Papaya
Pawpaw (Carica papaya, family Caricaceae, Order Brassicales, commonly called papaya in other countries) are highly productive fruit trees, native to meso-America and now grown in tropical and sub-tropical areas worldwide. They are enjoyed for their pleasant flavours and aromas, and also greatly valued as a rich source of vitamin A which is a major deficiency problem in much of the developing world. But for almost all the time they have been cultivated, growers have had to deal with their strange sexual behaviour which influences all the parameters of fruit production, without any real understanding of what’s going on and how best to optimise outcomes.
About 90% of all the flowering plants (angiosperms) are hermaphrodite or bisexual, and are normally self-fertile. Another 5% are monoecious, and most of the remainder are dioecious. A very small minority such as papaya are trioecious (sub-dioecious), with separate male, female and hermaphrodite plants. Seed germination is the principal way in which papaya are propagated and traditionally it has not been possible to determine whether a given plant will be male, female or hermaphrodite until it flowers. As only the latter two produce good quality marketable fruit, this results in much wasted time and resources raising and then eliminating plants. Only one in 10-20 plants need be male to ensure sufficient pollination of females, and depending on the climate, one or other of the bisexual or female plants may be preferred for fruit production. Bisexual plants are usually preferred in tropical climates and dioecious in sub-tropical, and this preference may mean other young plants are sacrificed. Traditional growers were also aware that varying environmental conditions each season, such as temperature, precipitation, nitrogen fertilisation, plant age, light intensity, photoperiod, mite infestation and mechanical injury (eg leaf shredding after storms or loss of storage tissue from pruning) could cause strange outcomes, with males sometimes producing fruits, females doing the equivalent of the virgin birth with fruit production in the absence of fertilisation (parthenocarpy), females temporarily becoming sterile or having long peduncles, trees producing normal-shaped fruit one year and deformed fruit the next, and individual trees changing to gynomonoecious or andromonoecious flowering.
Although a prominent feature of papaya behaviour, sex lability is not unique to this species and the phenomenon has been observed in more than 50 species across 25 families including such commonly known plants as asparagus, cannabis, castor oil, corn, cucumber, holly, orchids, spinach, summer squash, weeping willow, wheat, white mulberry and wild grapes. Usually hermaphrodites are more labile than the other sex forms. Dioecious plants under stressful conditions may convert to hermaphrodites or the opposite sex, and monoecious plants may change the ratio of male and female flowers. These changes are usually consistent with evolutionary adaptations that maximise reproductive success under difficult conditions, as the stress on females carrying fruits through to maturity is much greater than males producing pollen for only a limited time. Generally, stress of one form or another results in conversion from female forms to male, and optimal conditions the reverse.
Substantial advances in understanding the papaya story only began with systematic study in the early 1900s. Unravelling the mechanisms controlling sexual expression illustrates how scientific knowledge progresses and also how fundamental interpretations of data are always dependent on techniques available in different eras. While the story is still incomplete, many key features are now firmly established. Some of the major steps along the way were:
- In the 1930-40s breeding experiments of the different sex types showed that offspring occurred in the following ratios:
male X male – 1 female, 2 males,
female X male – 1 female, 1 male,
bisexual X bisexual – 1 female,
2 bisexual,
female X bisexual – 1 female,
1 bisexual, and
bisexual X male – 1 female, 1 male and 1 bisexual.
- These non-classical ratios provided support for the existence of a sex-determining gene that had three different forms (alleles) – M from male trees, Mh from hermaphrodite and m from female. These studies also suggested that male and hermaphrodite trees were heterozygous – Mm and Mhm resp, and females homozygous, mm. M and Mh were assumed dominant and m recessive, and dominant combinations MM, MhMh and MMh were lethal. This interpretation formed the basis of most theories over the next several decades, but it had to be successively embellished as new data came to hand.
- Flowers determining sex were found to be highly variable (more than 30 types), particularly amongst hermaphrodites, and in the 1950s it was suggested they could broadly be classified into 8 basic types, with (i) and (ii) being male type, (iii)-(vii) hermaphrodite and (viii) female. The different types had the following characteristics:
- staminate, – unisexual male flower on long peduncles, 10 stamens present in 2 whorls of 5 each
- teratological staminate – found on sex-reversing males, with some degree of carpel initiation and vestigial carpels (hair-like processes) at the base, can fruit in cooler weather
- reduced elongata – modified elongata flower, aborted pistil and reduced carpel size, more frequent in warmer conditions.
- elongata – refers to the shape of the pistil, develops into pyriform or cylindrical fruit, 5 lateral-fused carpels, and petals fused two-thirds length
- carpelloid elongata – transformation of the inner 5 stamens into carpel-like structures. Many sub-types depending on how many stamens are carpelloid, misshapen fruit
- pentandria – normal hermaphrodite, stepwise transformation of stamens to carpels, short corolla tube, 5 stamens in the outer whorl on long filaments, globose and furrowed pistil with 5-10 carpels
- carpelloid pentandria – all 5 stamens in the outer whorl become carpelloid, producing carpellodic fruit, especially under cool conditions, original carpels abort and flowers resemble pistillate type
- pistillate – unisexual flowers, larger than hermaphrodites, 5-carpellate, lack of stamens and the most environmentally stable of all types.
- In the 1940-60s a gene balance hypothesis proposed that the M and Mh alleles were on normal chromosomes (autosomes) and the m allele on ‘sex chromosomes’, although at that time there was no supporting cytological evidence of these. The M and Mh regions on the autosomes were degenerate, missing genes that were necessary for plant development. This model explained why homozygous dominant MM, MMh and MhMh were lethal, as heterozygous plants with at least one copy of m were viable.
- Also in the 1960s it was suggested that rather than a single gene controlling sexual expression, a number of these genes resided in a small region of the ‘sex chromosome’. This theory suggested there were 5 genes involved and various combinations of them produced the highly variable observed behaviour. It was also suggested the existence of hermaphrodite papaya could have been due to human selection ie it was a relatively recent development in the species. A real problem was that, like other theories on papaya, there was no definitive means of testing them.
- Another proposal in the 1960s was that papaya sex determination was of the XX-XY type that we’re familiar with in humans and most other mammals; namely homozygous XX being female and heterozygous XY being male. It was further posited that the Y chromosome has a region containing a lethal factor, and that Mh was a modified form of M while still including the lethal factor.
- In the 1990s a dominant male allele SEX1-M was proposed that promotes stamen development but suppresses that of the carpels ie it is a masculinizing factor. The dominant hermaphrodite SEX1-H allele was suggested to be an intermediate with the ability to induce stamens but only partially suppress carpels. The recessive female allele sex1-f promotes carpel development but is a null allele in terms of inducing stamens. Functional stamens and carpels would develop in heterozygous SEX1-H/SEX1-f plants, and the lethal factor linked to the SEX1-M and SEX1-H alleles would explain why homozygous dominant alleles do not survive and heterozygotes are viable.
- Finally in the last 15-20 years, numerous molecular biological studies have provided unequivocal answers to a number of these perplexing observations, interpretations and management issues. Up to the present, none of the vegetative papaya features (eg, stem and flower colour) suggested for their ability to discriminate sex type before flowering is reliable, and growers still have to put several seedlings in each planting hole and wait for flowering to know which can be culled. Modern DNA techniques can discriminate sex type well before this, but currently are only feasible with laboratory backup. This could change in the not too distant future as simple, cheap diagnostic genetic tests become available.
Papaya have 9 chromosome pairs and a small genome of 442 million base pairs with approximately 24,000 genes. The gene count may seem large but in fact is made up of many repeats, non-functional pseudogenes and nonexpressed DNA. It has been estimated that the minimum gene count for viability in Angiosperms is about 13,000. Approximately 96% of the genome has now been sequenced and mapped. The sex determining region is on an incipient Y chromosome which could not be differentiated from others using classical techniques. Separation began about 7 million years ago and the male specific region (MSY) is only about 10% of Y compared to 95% in humans with more mature chromosome divergence (240-320 million years ago). For such evolution to occur, it had to become non-recombinant to allow the MSY to develop and build up differences from the X. Regions just outside the MSY are changing about 7 times faster than the genome average, ie they are ‘hot spots’ responsible for the rapid changes that have occurred in this relatively short time. The corresponding MSY region on the X chromosome is changing much slower. Non-recombinant changes have led to degeneration and expansion in the MSY, mainly through insertions that occurred in 2 major evolutionary events, with loss of some key genes necessary for survival while still being present on the matching X chromosome; dominant homozygotes are thus lethal and heterozygotes viable. In breeding experiments the MSY behaves like a single (linked) genetic unit since it’s all on one chromosome and there’s no recombination with X. The hermaphrodite sex chromosome has an equivalent region (HSY) to MSY that diverged from it only 73,000 years ago, an incredibly short time in evolutionary history but still probably too early to be the result of human selection. It consists of 8.1 million bases compared to 3.5Mb on the matching X region, and it codes for 72 genes compared to 84 on the corresponding X chromosome region; 50 of these genes are common to both and there are 24 and 14 pseudogenes in the HSY and X regions resp. To date, papaya is the only plant species with fully sequenced and annotated sex chromosomes. The small papaya genome is probably the reason for genetic diversity between different varieties being relatively minor (correlation coefficient about 0.9) compared to other flowering plants. Self-pollinated hermaphrodites (cleistogamy) are just as variable as open-pollinated dioecious lines, seemingly from rare but still finite cross-pollination.
Two genes are likely involved as the initiators of sex determination, one a carpel suppressor or masculinizing gene for carpel abortion in male flowers, and the other a stamen suppressor or feminizing gene. These two genes operate in different time frames – abortion of stamens occurs before initiation of stamen primordia whereas the male sex determination gene aborts carpels at later developmental stages. Remnants of the aborted gynoecium are a feature of the male flower structure, and with good growing conditions, a few male flowers may not undergo complete carpel abortion and can form fruit. Arabidopsis thaliana is the closest relative to papaya in the Brassicales that also has sex chromosomes, and this species has been extensively studied by plant geneticists. Many genome parallels exist between the two species that indicate the papaya female sex determining gene is likely to be an upstream regulator of 2 genes called APETALA3 (AP3) and PISTILLATA (PI) that cause early abortion of stamens. A thaliana also produces a gene called ATA1 that is expressed only in male flowers during development and is homologous to similar genes found in corn and white campion. ATA1 is associated with normal pollen formation and although it’s probably also involved in the masculinizing pathway in papaya it might not be the initial gene that sets the whole flower development cascade underway. Approximately 180 genes are thought to be involved in producing fully functional flowers.
At least 2 genes differentiate the M and Mh chromosomes; one controls the long peduncle on male trees and the other is a masculinizing gene that controls carpel abortion in male flowers. As embryo abortion occurs 25-50 days after pollination, there is a regulatory gene that’s essential to early embryo development that resides in this region and has degenerated on both the M and Mh chromosomes but is still functional in the X region; this is the cause of homozygous dominant allele lethality. It has been reported that one of the key genes in papaya sex differentiation between males and hermaphrodites and not found on the X is similar to one found in A thaliana called SHORT VEGETATIVE PHASE (SVP) . SVP has been shown to initiate flower development, and in papaya it is functional in the MSY but not in the HSY. Thus it would seem that in males, SVP sets in train carpel abortion whereas in hermaphrodites a varying number of carpels remain functional and can therefore be fertilised and bear fruit. This whole field is advancing rapidly and the hope is that it won’t be too long before all the key sex determining genes are identified and characterised. The evidence so far is that these few genes are the initial triggers for setting in train male, hermaphrodite or female development through subsequent genetic, epigenetic and plant hormone control.
Some examples of other practical developments and possibilities that have followed this genetic work include
- Cloning of a single gene that codes for an enzyme called lycopene beta-cyclase which determines red fruit flesh colour. In many countries red flesh papaya are preferred over the more usual yellow/orange types and accordingly fetch higher prices. The allele for red colour, which produces a non-functional enzyme, is recessive, and this is why typically with open pollination this feature can be quickly lost and fruit revert to the less strongly coloured forms. Healthy pro-vitamin A carotenoids in red flesh fruit consist mainly of lycopene whereas in the yellow/orange forms with the dominant allele, almost all lycopene is enzymatically converted to other carotenoid forms.
- The accelerated impetus for genetic work on papaya stemmed in large part from the disease Papaya Ring Spot Virus (PRSV). This is a serious problem in all papaya growing regions worldwide and can devastate whole orchards. In Hawaii, papaya had been one of the top 3 fruit crops and they were threatened with total collapse of the industry because of the spread of the virus across all the islands. C. papaya is the only species in the genus and with a small genome there were limited opportunities to use classical breeding strategies to develop more resistance – none was sufficiently successful. Following sequencing of the PRSV and identification of the viral resistance gene that produced a coat protein, a major collaborative research effort was undertaken to develop a transgenic solution by inserting this gene into papaya. Sunup (red flesh) and Rainbow (yellow flesh) became the first such tree crop fruit to be approved for general release in the US and local growers quickly moved to incorporate them in production; subsequent approval in foreign markets such as Japan cemented their place as a valuable commercial crop and saved the industry. Other major papaya producers in Latin America and South East Asia have found PRSV infections in their regions are due to different strains, and with assistance and a technology transfer program are developing their own transgenic solutions.
- As genes are characterised, particularly those in the MSY and HSY, and their role in plant and fruit development clarified, there is the prospect of having pure breeding lines that avoid the multi-planting problem associated with conventional seedling propagation. As a number of other species in the Brassicales have already been cloned and have larger genomes with many more genes whose purpose has been identified, parallel genes can be seen in the C. papaya genome that regulate features such as precocity, stem height, photoperiod response, climacteric behaviour, fruit levels of sugar, starch, phytonutrients and aromatic volatiles, cellulose synthesis for tree strength, fruit firmness and size, storage properties and disease resistance. Exploitation of this expanding collective knowledge will support improved economic production of superior fruit with greater control and/or predictability under varying environmental conditions.
Sexing Papaya: what have I got in the backyard?
Domestic and commercial papaya growers world-wide almost universally use seedlings for papaya propagation, mainly because seed supply is plentiful and cheap, germination is easy and the juvenility period is so short – 9 months under ideal conditions. But with seedlings you have to wait till flowering time to know which of the three main gender types a given plant will be ie male (staminate), hermaphrodite (bisexual) or female (pistillate). Modern genetic techniques can answer this question much sooner, but without research lab facilities the rest of us have to wait for events to unfold. The dioecious types perform better in sub-tropical climates like ours, and only one male is required to pollinate several females; fruit can be formed parthenocarpically but quality is compromised. So given you’ve probably sourced your seed from open-pollinated dioecious or self-pollinating hermaphrodite plants, what proportion of each gender have you grown up and which ones are either in excess or unwanted and can be culled? Obviously the sooner you’re able to answer this question following flowering and then continue with only the desired types, the better. First you should read the companion article in this series, ‘The mysterious antics of papaya’, to allow some understanding of the markedly variable behaviour and sexual properties of the species. Then when you have some feel for the complexity of what you’re dealing with, the intention of this article is to move on to practical identification of the various types. Of the three main genders, hermaphrodites show the greatest variation in features, and their behaviour is also the most influenced by cultural and environmental factors.


For female and hermaphrodite plants this is short, usually with a single axial flower but sometimes up to 2-3, occurring as short internode cymes. Ideally the peduncle should be 4-6cm long so that when fruit mature they’re not all tightly jammed up against one another on the trunk, as this leads to misshapen fruit and difficulty in harvesting. In males, there may be many dozens of flowers on drooping peduncles 60-90cm long with flowers arranged as long internode cymes (raceme-like).
Flower morphology – exterior
- Staminate. The small narrow flowers have a fused tubular 5-lobed corolla that extends almost the whole flower length. There are 5 separate petals at the throat of the corolla, with stamens in the centre partly visible. They are the smallest of the 3 main types.
- Pistillate. Globular-obovoid with separate twisted petals inserted at the base of the slightly inferior ovary, with 5 apical stigmatic lobes protruding. Largest of the three types.
- Hermaphrodite types. These are intermediate in size between the other 2 types, and the apical stigma is apparent in all cases.
- Elongata. This is the normal hermaphrodite, so named because of the elongated pistil that leads to more elongated cylindrical fruit. Petals are not twisted and are fused together for a proportion of their length to form a corolla tube free from the pistil. In the unopened bud, there is usually a slight restriction around the throat region.
- Pentandria. Globular-elongated. Petals are free for most of their length but more fused to each other and to the pistil base than in pistillate flowers.
- Carpelloid forms. The least satisfactory of the hermaphrodites and the most variable. Petals can be inserted at the pistil base or fused for up to 70% of their length




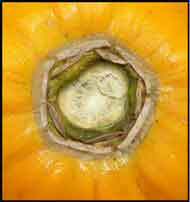
Flower morphology – interior
Gently peel off the petals one at a time as needed. You might find a hand lens helps you see some of the following details.
- Staminate. The 10 stamens can now be seen more clearly in 2 separate alternating whorls of 5, with one series being sub-sessile anti-petalous and the other anti-sepalous on short filaments. There is only a rudimentary and non-functional central pistil (pistillode).
- Pistillate. There are no stamens. The 5-carpellate pistil is usually smoothly circular or slightly undulate, and the apical stag-like stigma is now clearly visible.
- Hermaphrodite types.
- Elongata. This is the preferred type of hermaphrodite. There are 10 stamens at the throat of the corolla tube. The stamens are arranged similarly to staminate flowers. In some plants the anthers and stigmas are at the same level and in others different, with consequent effects on ease of pollination. The pistil is 5-carpellate but can vary from 1-10.
- Pentandria. So-called because it has 5 fairly short anti-sepalous stamens. These are fused to both the pistil and the petals at the base, which is just below the throat of the short corolla tube. The pistil is 5-carpellate and deeply furrowed with the stamens lying along these grooves.
- Carpelloid forms. Carpellody is where stamens lose their pollen forming ability and become carpel like by fusing with the pistil. There is great variation in these forms. Petals may be almost free from one another or fused with others for up to 70% of their length, even within a single flower. There may be 2-10 stamens in different stages of transition from anthers to carpels, and their fusion with petals and pistil can also vary. Pistils are often distorted with 5-10 carpels. Sometimes there may be faulty fusion leaving ovules exposed and in others the original 5 carpels may abort. Many of these unsatisfactory variants are driven not only by the genetic make-up of the plant but by un-favourable cultural and environmental factors.
Fruit
You can firm up your diagnosis when the plant begins to fruit.



- Pistillate. Globose-obovoid without furrowing. Surrounding the stem at the base of the fruit, there are 5 separate scars where the 5 individual petals have abscised. This type is the least subject to cultural and environmental influences.
- Hermaphrodites.
- Elongata. The best fruit of the 3 hermaphrodite types. The most common form is 5-carpellate and pyriform or cylindrical. Flowers with only a few carpels form narrow fruits similar to bananas.
- Pentandria. Sightly oblong-cylindrical with 5 longitudinal furrows. The fruit stem end shows a continuous circular scar which comes from the remnants of the abscised corolla tube.
- Carpelloid forms. The least satisfactory of the 3 basic hermaphrodite types. They are usually misshapen, sometimes grotesquely, and are not commercially marketable. When you cut the fruit you will sometimes see more than a single, central seed cavity. Even for the home grower it’s probably not worth keeping the tree.


Sex-reversing males (teratological staminate flower type)
With adverse cultural and environmental conditions (eg temperatures below 12°C through our winters) these trees which normally produce male flowers can become hermaphrodite. Incidentally, although commonly used in the literature, the term ‘sex-reversal’ is somewhat misleading. True reversal would take male through to female, but in these plants their gender change only goes half-way, having both male and female organs The flowers are larger and more bulbous than pure male types. They usually occur terminally on the peduncle and have an elongated pistil with a variable number of surrounding stamens. The fruit are narrow, elongated, furrowed and sometimes quite distorted. Certainly not commercially marketable and even for the home grower, not worth producing. If you decide to keep the tree, at least remove the fruit to prevent waste of plant resources.


Grafting Papaya
Isolated published scientific studies report it’s possible although difficult to graft papaya. But you probably won’t see any mention of this propagation technique in your usual reference sources because of several important characteristics of the species.
- Almost exclusively, growers from backyarders to big commercial operations worldwide use seed propagation because each fruit produces hundreds of viable seeds that store well, germinate easily and can produce fruit in less than a year with ideal climatic conditions and optimal management.
- It is one of the very few major fruit tree crops in the world propagated in this way, as usually there’s a more prolonged juvenile period over some years with seedlings, and vegetative propagation usually halves this non-productive period.
- The superior fruiting qualities of selected cultivars, normally desired in other fruits which require vegetative or micro-propagation techniques to be maintained, are less important in papaya. The hermaphrodites favoured in tropical climates can become almost pure breeding lines after several generations of controlled in-breeding as they primarily self-pollinate. In sub-tropical climates like ours, the dioecious varieties perform better, and although subject to normal sexual variation can be manageable with selection of seed from superior trees and by restricting open pollination from unknown males or hermaphrodites. At least half the gene pool in this situation (the female side) has been selected.
- Papaya have very strong apical dominance which results in them normally being single-trunked, with axial fruiting occurring as meristematic buds located in the tip region differentiate into flowers. As an herbaceous, soft wooded and hollow-stemmed species, this region is very delicate and not hardened to minimise desiccation as in other more woody species. Lateral buds lower down the stem remain inhibited under normal conditions (paradormancy), but can develop slowly following injury or pruning of the apical region. This means scion material for grafting is typically non-ideal, either too delicate from the tip and requiring fastidious control of humidity, or strongly inhibited if lower segments are used.
- Normal grafting (eg whip or cleft) requires matching of the cambial regions of scion and rootstocks, but unless relatively mature males are used for the rootstock there’ll be a gross mismatch in diameters between them.
- The species is laticiferous, which interferes with timely union of cambial regions from scion and rootstock.
A principal problem that all growers face with seedling propagation is that it’s not possible to determine sex in the field before flowering. As a consequence the common practice is to put 3-4 seedlings in the one planting hole, wait till flowering then cull those males or non-vigorous females not needed. With dioecious varieties only one male is needed to pollinate several females and this means considerable time and resources have been wasted raising these superfluous plants. For the home grower who is only interested in a small number of plants, Murphy’s Law inevitably kicks in and the several potted plants you’ve grown up ready for in-ground planting could be almost all males. When propagating these plants by seeds you should replace your chosen 3-4 mature females every few years to keep up yield and quality – always have new plants coming through. Usual practice is to kill the unwanted excess males and hope the females give good quality fruit – but it’s still a bit hit and miss.
It is possible to graft – unwanted males would have been sacrificed anyway and if the graft doesn’t take there’d be nothing lost. For example, a female might be in the last year of production and had been topped to reduce height. Expected lower branching would have developed, so providing smaller diameter scion material in vegetative growth mode. A decision then has to be made on which way to go, the tip region or lower down the branch? If you decide on the latter, you would be hoping the graft would take with sufficient lateral bud growth before it all dried out. This means it’s far easier to match diameters of scion and rootstock for a cleft graft as the small male seedling has just started to produce flowers. The first picture show it can work & you could be confident of getting fruit the equivalent of your best quality tree; it could become the main way to generate stock for each renewal cycle. There’s also a further favourable outcome – males are usually more vigorous than females so a grafted plant on male rootstock could be more vigorous, giving greater yield. A tip if you decide to have a go – the very soft stem material provides little mechanical strength to hold the scion firmly, so bind tightly with grafting tape and keep the scion relatively short so it won’t be too top heavy.

Update
Literature reports indicate cleft grafting can be successful with papaya, and consequently this might be the first technique Iyou try. However the very soft-wooded lateral branches that are the most likely source of suitably responsive dormant buds mean that without external support, the slowly desiccating scion can wilt and become difficult to maintain in firm and fixed cambial contact with the rootstock in the period before the union becomes functional. A side graft could effectively address this limitation as the remaining root stock above the graft area would provide a ready-made stabilising ‘post’ for anchoring the scion. A further consideration is that root stock leaves left above the graft would ensure continuation of sap flow around the graft region and enhance the chances of take. Within two weeks you might get new growth in the scion, which can rapidly progress to the level shown in the picture. It could be a better way to go than top working with a cleft graft.

Papaya side graft update 2
This picture taken several months later shows the plant growing on nicely after not doing very much over an unusually long, cold winter and spring that year. Flower buds have already formed and hopefully good fruit would be expected to come through in the not-too-distant future given the female scion pedigree.

Papaya side graft update 3
Fruit set was in autumn, growth stalled through winter, and the picture shows fruit afterwards maturing nicely. This illustrates such vegetative propagation gives fairly rapid fruit production of selected phenotypes. The graft region can be seen at about knee height.

s