The wondrous ways of angiosperm reproduction
There are more than 350,000 angiosperm species worldwide (the flowering plants), and many of these include fruit trees we’re interested in. They usually reproduce sexually, with production of fruit containing seeds which protect embryos from desiccation, predation or disease till they are mature enough to be dispersed in climatic conditions favourable for subsequent germination and seedling growth. However a small minority can produce seeds asexually, where the normal sexual steps of meiosis (see definitions below) and fertilisation are bypassed. There are a number of intricate pathways in which this can occur as described below, collectively known as apomixis. It’s quite unevenly widespread having co-evolved frequently, and is found in more than 80 families and 400 species; these numbers continue to grow as appropriately designed studies are undertaken. Another way that plants can reproduce is vegetatively, eg by stolons and rhizomes, but this is a separate topic and will not be considered in the following commentary.
We’re all familiar with particular fruit tree seedlings that come through fairly true to type when the norm is to find that properties can vary significantly from their parent/s (sexual variation). It can happen in several ways, eg self-fertilisation with less genetic difference between sperm and ovule, small genome size in the species, or comparatively low genetic heterogeneity in the species, but the possibility of apomixis underlying the phenomenon should be kept in mind. To understand what this is in angiosperms, it’s best to first go through the more common process of sexual reproduction, focussing on the female line where most of the action takes place. Sexual reproduction leads to mixing of genes and hybrid vigour that can increase the chances of species survival when environmental conditions, opportunities and threats may vary over time. Consequently it’s not surprising to find that evolution has resulted in all sorts of mechanisms to favour sexual cross-pollination, eg dichogamy and herkogamy. In contrast, apomictic reproduction leads to offspring being (mainly) clones of the maternal parent. These may be highly desirable because they can fix important plant and fruit properties such as vigour, disease resistance, precocity, fruit size, yield and quality. The alternative for us to achieve this predictability is to use vegetative propagation techniques such as cuttings and grafting that all require time, costs and expertise. Apomixis is usually facultative rather than obligate, ie pollination with sexual fertilisation may be the principal pathway for plant reproduction, but there is a safety mechanism to ensure survival if plants are too far removed from others or pollination vectors are inadequate. It is also commonly associated with hybridisation, where different parent genomes can lead to chromosomal pairing problems, asynchrony in gene expression, timing and interaction that together can cause meiotic failure. Polyploidy is another common feature of apomixis; this has similar effects on gene expression and hormone regulation, with loss of self-incompatibility that normally promotes cross-pollination, making it possible for some plants to self-reproduce clonally.
The definitions and diagrams below have been adapted from Hörandl, Classification of Asexual Organisms, Taxon (2018) 67, 1066-1081.
Definition of Terms:
Adventitious embryony: see sporophytic apomixis.
Agamospermy: Reproduction via asexually formed seed. Synonym of apomixis.
Apospory: Development of an unreduced female gametophyte out of an unreduced initially adventitious somatic cell.
BII offspring: Sexually formed offspring of a facultative apomict, i.e., the embryo sac is formed meiotically, and the reduced egg cell is fertilized (n + n).
BIII offspring: The embryo sac is formed apomictically (either via Diplospory or Apospory). The unreduced egg cell is fertilized, resulting in a ploidy increase in the embryo (2n + n).
Diplospory: Development of an unreduced female gametophyte out of an unreduced megaspore.
Facultative apomixis: A single plant can produce sexual and apomictic seeds within the same generation. The resulting offspring have the same ploidy as the mother plant, but are partly clonal (2n + 0), and partly recombined (n + n). Often this term also includes cases of Partial apomixis (see below).
Haploid parthenogenesis: The embryo sac is formed sexually (i.e. after meiosis). The reduced egg cell is not fertilized, but develops parthenogenetically. The embryo has half the ploidy of the mother plant.
Parthenogenesis: The development of an egg cell into an embryo without fertilization.
Partial apomixis: Occurs when embryo sac formation is not coupled to parthenogenesis and results in ploidy shifts in the embryo. Sometimes subsumed under Facultative apomixis. For the increase of ploidy (2n + n) see BIII offspring, for the decrease of ploidy (n + 0) see Haploid parthenogenesis.
Polyembryony: Two or more embryos develop within one seed; this can result from Sporophytic apomixis (see below), when both sexual and apomictic embryos are formed.
Pseudogamy: Pollen-dependent apomixis; the egg cell develops without fertilization, but the polar nuclei are fertilized by one or two sperm nuclei. This is often essential for endosperm development and formation of functional seed.
Sporophytic apomixis: Development of an embryo directly from a somatic cell of the ovule (= adventitious embryony). The embryo is a clone of the mother plant (2n + 0), and may also develop in parallel with the formation of sexual embryos (n + n).
Additional Terminology:
Antipodals: cells, commonly three in number as in an eight-nucleate embryo sac, located at the other end of the embryo sac from the female gamete
Apomixis: plant asexual reproduction through seed
Central cell: the large cell initially usually containing two nuclei (polar nuclei) that makes up the bulk of the eight nucleate embryo sac
Dichogamous: separation in flowering time of anther dehiscence and stigma receptivity in the one flower
Diploid: plant cell with two basic haploid sets of chromosomes in the nucleus gametophyte: phase of a plant’s life cycle that bears gametes, usually after mitotic events
Meiosis: the two-stage division of a diploid nucleus during which gene recombination occurs and the number of chromosomes is halved
Mitosis: the normal division of a nucleus, whether haploid or diploid, during which chromosomes are replicated
Haploid: plant cell having a single set of chromosomes in the nucleus
Herkogamy: where pollen presentation and receipt is spatially separated within flowers
Meristem: region of a plant where undifferentiated cells divide, generating new cells that ultimately differentiate
Rhizome: a slender to much swollen underground stem that grows more or less horizontally
Stolon: prostrate or trailing more or less above-ground stem which produces roots and sometimes erect shoots at its nodes
Synergids: part of the egg apparatus. Two cells at the apex of the embryo sac that are closely associated with the egg cell and directly involved with the process of fertilisation
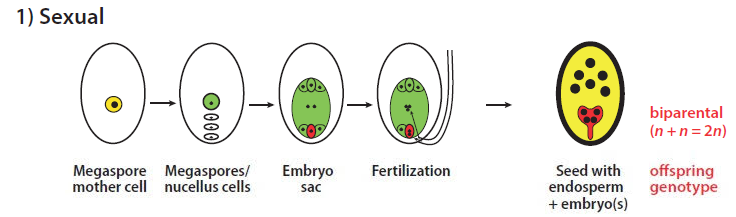
Sexual reproduction begins when plant meristematic buds switch from vegetative to reproductive mode. This leads to development of flowers instead of leaves, and formation of male (anther) and female (ovule) reproductive organs. Many diploid cells differentiate in anther pollen sacs and form microspore mother cells, but in ovules which ultimately form the seeds, only a single cell called the megaspore mother cell differentiates in each. Both of these types of precursor cell undergo 2 stage meiosis, where the diploid nucleus with 2n sets of chromosomes is halved, chromosomal exchange can occur, and 4 haploid cells with n chromosomes are most commonly formed (megaspores). The male microspores, also with n chromosomes, develop into pollen grains, whereas in the ovules, each megaspore mother cell most commonly forms 4 megaspores, 3 of which degenerate, leaving the largest one as the functional megaspore. Each megaspore then undergoes 3 mitotic divisions to produce 8 haploid nuclei, initially in a single cell structure that subsequently produces a 7-celled embryo sac – at one end one there is an egg cell (n, red) and 2 synergids (n), 2 form the central cell (2n), and at the other end 3 form the antipodal cells (n). With efficient and genetically compatible fertilisation there is deposition of pollen grains on the flower stigma, pollen germination, and then growth of a pollen tube containing 2 sperm cells towards the ovule, 1 of which fuses with the egg cell to form a diploid embryo (biparental above, giving 2n chromosomes), and the other to the central cell that forms the triploid endosperm (3n) which serves as an early nutrient supply for seed germination and early growth. The ovule tissues surrounding the developing embryo and endosperm form the seed coat.
There are 5 main variant apomictic mechanisms with possible interactions between them considered below. They differ from sexual reproduction principally in 3 ways, (i) the absence of meiosis, (ii) the possible absence of pollination (parthenocarpy), and (iii) the central cell can form the endosperm either with (pseudogamy) or without fertilisation (autonomously). In some cases apomixis can occur together with sexual reproduction, and well-known examples amongst fruiting species are mango and citrus where in a single seed there may be one sexual embryo and one or more maternally-derived asexual ones. Some viable pollen production usually persists in facultative apomictic plants and can be involved in central cell and egg cell fertilisation, eg with pseudogamy, BIII hybrids and adventitious embryony. Diplospory or apospory can occur in different species within a genus, varieties within a species, in a single plant and in single ovules, with the relative proportions of the two being influenced by genetics and environmental conditions.
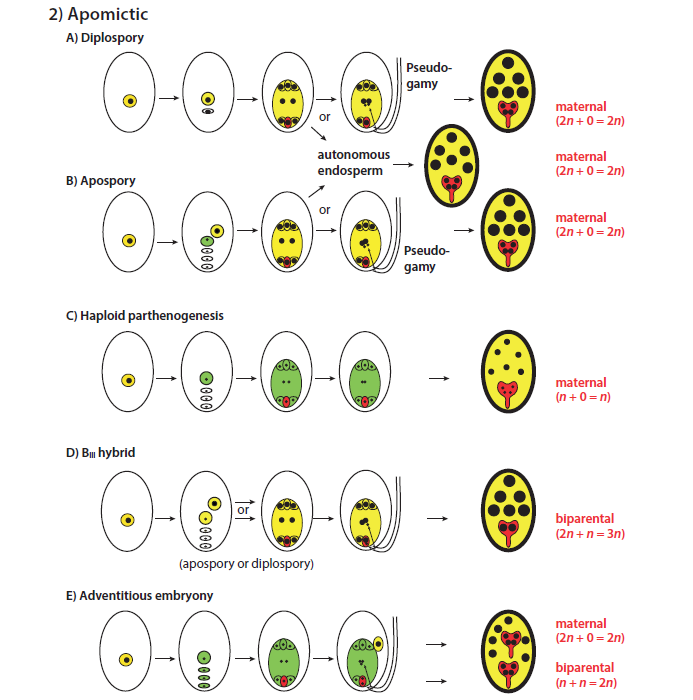
Diplospory (A above)
Here we have a megaspore mother cell that doesn’t undergo meiosis and remains diploid, while still progressing through the usual subsequent stages to form an embryo which in this case is entirely derived from the diploid maternal parent – hence the term diplospory, with 2n + 0 indicating zero nuclear chromosome sets come from a sperm cell. Double fertilisation may (i) or may not (ii) occur. In case (i, pseudogamy), the central cell (4n) is fertilised by both sperm nuclei (2 X n) and becomes hexaploid, maintaining the normal 2:1 maternal/paternal chromosomal ratio; however this ratio is not always seen in different species. Self-incompatibility that normally encourages cross-pollination needs to have been lost if central cell fertilisation is to occur by selfing or by a separate plant if a clone. Pseudogamy seems to be necessary for subsequent seeds to be able to produce at least some viable pollen production. In case (ii), commonly the central cell may remain unfertilised but nevertheless continue to develop autonomously. Rubus, sub-genus Rubus (brambles, blackberries) is endemic to Europe, and of the 750 species accepted there, only 4 are diploid and these exhibit obligate sexual reproduction. In higher ploidy levels, obligate diplospory is seen with triploids and facultative apomixis with tetraploids. Both apospory and diplospory are seen in some species. Cherry guava is another example of a facultative diplosporic apomict.
Apospory (B)
This term implies that a megaspore is derived from a diploid somatic cell and all 4 of the megaspore mother cellderived mother cells degenerate. The somatic cell progresses through subsequent developmental stages, commonly with double fertilisation (pseudogamy) and rarely without the egg cell being fertilised (autonomously). The result in either case is a diploid embryo formed from the maternal line with zero male chromosomes contributed by fertilisation (2n + 0). Medlar is an example of a facultative pseudogamous aposporic apomict.
Haploid Parthenogenesis (C)
Rarely a normal 7-celled megaspore-derived embryo sac is formed but because of some disruption of the normal fertilisation step, the egg cell continues to develop parthenogenically into an embryo, even though only haploid (n + 0).
Blll Hybrid (D)
In this case the megaspore mother cell remains diploid and either the functional megaspore (diplospory) or a somatic cell (nucellar or integument, apospory) undergoes mitosis producing the 7-celled embryo sac which is then double fertilised. The result is a triploid embryo (2n + n).
Adventitious embryony (Sporophytic apomixis, E)
This is the most common form of apomixis, particularly in tropical and sub-tropical species. It usually occurs later in ovule development than gemetophytic apomixis. Normal meiosis and mitosis take place with formation of a haploid egg cell and 7-celled embryo sac, which is then fertilised forming a sexually derived embryo (n + n). But in addition, one or more adventitious diploid cells also may enter the embryogenic line and develop parthenocarpically, so that there may also be asexually derived embryos (2n + 0). Double fertilisation is necessary for formation of the sexual embryo and endosperm that nourishes developing embryos. Once fertilised, the sexual embryo sometimes degenerates, leaving only the asexuals. Rose apple (Syzygium jambos) is an example of a polyembryonic species, and the proportion of asexual/sexual embryos can vary depending on varietal genetics and environmental conditions. This ratio in Citrus is commonly about 3:1 (but a range of 2-10 or more); in Eureka lemon and Dancy mandarin, asexual seed ranges from 33% to 100% respectively. Competition for endosperm nutrients can also influence these proportions.
All up a complicated story, and although much less frequent than sexual reproduction, apomixis occurs naturally across a wide range of flowering plants. Considerable research effort is going into the area, and molecular techniques are rapidly improving our understanding of the genetics and physiology involved, eg amongst about 30,000 genes in Citrus, those responsible for polyembryony have been narrowed down to a region containing only 11 genes. The hope is that in the not too distant future it will be possible to generate important species and varieties superior in different ways to those currently available.